ABSTRACT
- During the last two decades, the many developments in the treatment of movement disorders such as Parkinson disease and dystonia have enhanced our understanding on organization of the basal ganglia, and this knowledge has led to other advances in the field. According to many electrophysiological and anatomical findings, it is considered that motor information from different cortical areas is processed through several cortico-basal ganglia loops principally in a parallel fashion and somatotopy from each cortical area is also well preserved in each loop. Moreover, recent studies suggest that not only the parallel processing but also some convergence of information occur through the basal ganglia. Information from cortical areas whose functions are close to each other tends to converge in the basal ganglia. The cortico-basal ganglia loops should be comprehended more as a network rather than as separated subdivisions. However, the functions of this convergence still remain unknown. It is important even for clinical doctors to be well informed about this kind of current knowledge because some symptoms of movement disorders may be explained by disorganization of the information network in the basal ganglia.
-
Keywords: Basal ganglia; Motor information; Somatotopy; Movement disorders; Electrophysiology
In the early years of the 1990s, the basal ganglia circuitry was summarized in several parallel loops with the “direct and indirect pathway model.” After the publication of this model combined with spread of stereotactic surgeries for deep brain stimulation therapy of movement disorders such as Parkinson disease and dystonia, clinical and experimental interest in organization of the basal ganglia has revived. A large number of anatomical and electrophysiological findings on the basal ganglia network have supported this model. However, it may be oversimplified. Although many researchers have tried to explain the pathophysiology of movement disorders by firing rate changes through these two pathways, it is still difficult to understand the mechanisms of most of their symptoms.
Recent studies have proposed some additions to this model and insisted on their importance. With regard to the organization of several loops in the basal ganglia, some convergence of information from different cortical areas was proved. Concerning the details of the loops, the role the subthalamic nucleus (STN) as a part of the “hyperdirect pathway” has occupied the interest of many researchers. Besides, one of the topics on motor information in the basal ganglia is somatotopy.
In this article, I review the current knowledge on organization of motor information in the basal ganglia, with special emphasis on assessing the electrophysiological evidences.
Current Model of the Basal Ganglia and Its Electrophysiological Evidences
- The basal ganglia receives inputs from wide areas of the cerebral cortex, in particular, from the frontal lobe. The information processed in the basal ganglia is transmitted back primarily to the cerebral cortex, via the thalamus, to form a cortico-basal ganglia loop.1,2
- In this circuitry of the basal ganglia, the striatum receives direct excitatory cortical inputs and projects them to the output nuclei of the basal ganglia, i.e., the internal segment of the globus pallidus (GPi) and the substantia nigra pars reticulata (SNr), via two major projection systems, the “direct” and “indirect” pathways. The direct pathway arises from the gamma-aminobutyric acid (GABA)ergic striatal neurons that contain substance P and projects monosynaptically to the GPi/SNr. The indirect pathway arises from the GABAergic striatal neurons that contain enkephalin and projects polysynaptically to the GPi/SNr via the sequence of connections involving the external segment of the globus pallidus (GPe) and the STN. In addition, dopaminergic projections from the substantia nigra pars compacta (SNc) differentially modulate the activity of striatal projection neurons in the direct and indirect pathways. Dopamine excites striatal neurons in the direct pathway through dopamine D1 receptors (D1Rs), whereas it inhibits striatal neurons in the indirect pathway through dopamine D2 receptors (D2Rs).2,3 Moreover, recent studies have proposed the idea that the STN receives direct cortical inputs, and that the cortico-STN-GPi/SNr pathway, called the “hyperdirect” pathway, also considerably influences the GPi/SNr activity.
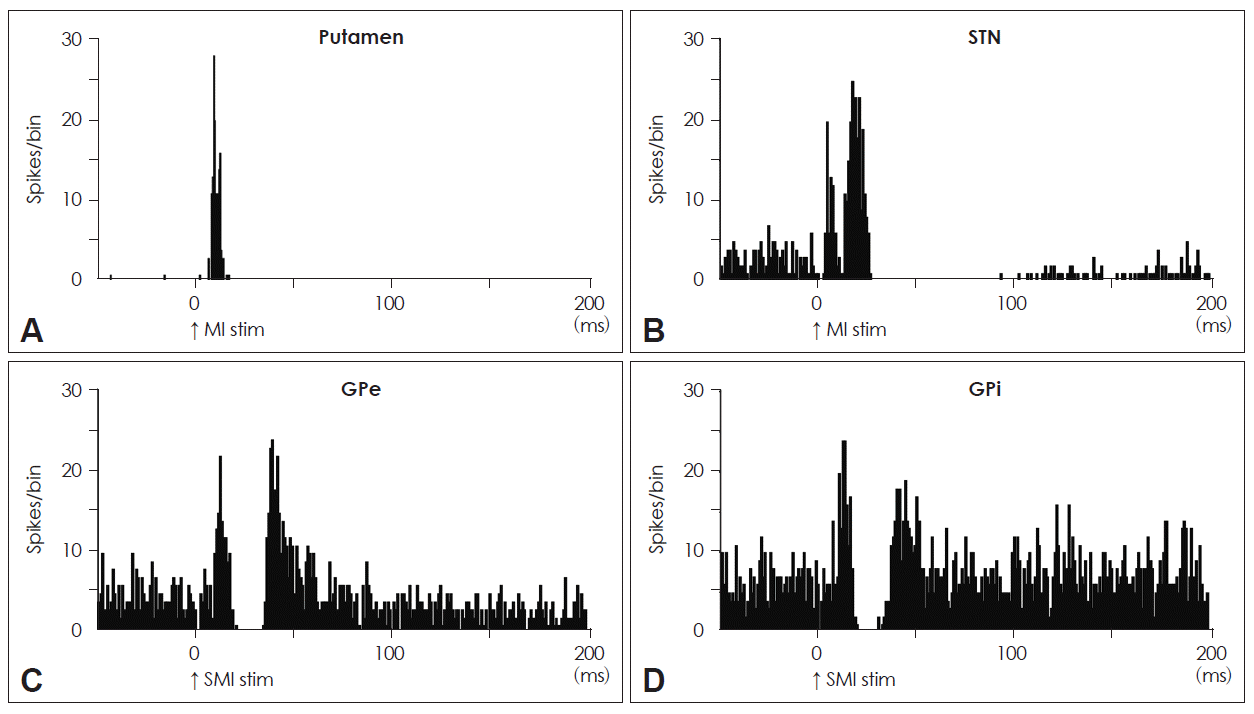
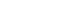
- This current model has been supported not only by many anatomical experiments4–10 but also by the following electrophysiological findings (Figure 1): the stimulation of the motor-related cortical areas evoked 1) orthodromic spike discharges in striatal neurons11 (Figure 1A); 2) induced a response composed of early and late excitations in STN neurons (Figure 1B)12,13; and 3) induced an early, short-latency excitation, followed by an inhibition and a late excitation in GPe (Figure 1C) and GPi (Figure 1D) neurons of monkeys.12,14–20 The early excitation evoked by the cortical stimulation of STN neurons is caused by the direct cortico-STN projections, and one of the main origins of the late excitation is considered to be the disinhibition in the STN through the cortico-striato-GPe-STN pathway. On the other hand, with regard to the responses of GPe and GPi neurons to cortical stimulation, the early excitation is derived from the cortico-STN-pallidal pathway, while the inhibition and late excitation are considered to be principally mediated by the cortico-striato-GPe/GPi and cortico-striato-GPe-STN-GPe/GPi pathways, respectively.
Parallel Organization of the Cortico-Basal Ganglia Loops
- The cortico-basal ganglia loops are composed of several parallel, segregated, and functionally distinct but homologous loops1,8 such as the motor, oculomotor, prefrontal, and limbic loops. Through these multiple loops, the basal ganglia control limb movements, eye movements, higher brain functions, and emotions. Accumulated evidence also suggests that each of these loops may consist of several subcircuits with single cortical areas of origin and permit highly specialized parallel processing of ample and various signals to be transmitted concurrently.1,2
- Despite this parallel organization, the cortico-basal ganglia loops should be comprehended more as a network rather than as subdivisions with rigorous boundaries. For example, in the case of the motor loop, although each input from various cortical areas, such as the primary motor cortex (MI), supplementary motor area (SMA), and premotor cortex (PM), is processed principally in a segregated manner, considerable convergence of the cortical inputs occurs in the basal ganglia. Some anatomical experiments showed that the projections from the MI, SMA, and PM partially overlap in the striatum,21,22 and an electrophysiological study revealed that around 20% of putamen neurons receive convergent inputs from the MI and SMA.11 Another experiment showed that around 30% of STN neurons receive convergent inputs from the MI and SMA.13 The functions of these convergences remain unknown. On the other hand, striatal neurons that receive inputs from the MI, SMA, and MI+SMA project to the segregated parts of the GPe and GPi,23 and it has been reported that the percentages of MI+SMA-receiving neurons in the GPe and GPi were still 20–50%.14 Thus, futher convergence does not seem to occur in the striato-GPe/GPi and the STN-GPe/GPi projections.
Somatotopic Organization of the Basal Ganglia
- Meanwhile, each nucleus of the basal ganglia shows somatotopy, and information from different types of somatotopy is well preserved through the cortico-basal ganglia loops. The most basic method to investigate this is to examine its fiber connections with other brain areas whose somatotopy is clearly identified. For example, anterograde tracers are injected into the orofacial, forelimb, and hindlimb areas of the MI, and then terminals in the striatum and STN are observed.5,21 Trans-synaptic anterograde and retrograde tracing allows the investigation of areas that are linked to the origin via synapses.7,9,10 Fiber connections can also be studied by electrophysiological methods. Stimulation of the MI induces responses in the corresponding areas in the striatum, STN, GPe, and GPi.11,13,14
- The striatum, an input station of the basal ganglia, receives excitatory inputs from all areas of the cerebral cortex, except for the primary visual cortex. The caudal region of the putamen, which is posterior to the anterior commissure, is considered to be the motor territory and shows clear somatotopy (Figure 2C).9,11,21 It has two sets of somatotopic representations in the lateral (MI domain) and medial (SMA domain) parts. The projections from the MI and those from the SMA converge in the medio-lateral central zone. In each somatotopic representation, the orofacial, forelimb, and hindlimb areas are arranged from ventral to dorsal.
- The STN, another input station of the basal ganglia, receives cortical inputs from the frontal lobe. The dorsal part of the STN is the motor territory and shows somatotopic organization (Figure 2A).5,13 The MI projects to the lateral part (MI domain) of the motor territory of the STN, whereas the SMA projects onto the medial part (SMA domain). The orofacial, forelimb, and hindlimb areas of the MI project onto the lateral to medial parts of the lateral STN, whereas those of the SMA project onto the medial to lateral parts of the medial STN. The MI also partly projects to the somatotopically corresponding body parts in the SMA domain, and the SMA partly projects to the MI domain.
- The motor territories of the GPe and GPi form the ventral two-thirds of the caudal GPe and GPi, which is the area where the motor territory of the striatum projects.24 The somatotopy in the GPe and GPi can be drawn by observing the responses evoked by the stimulation of the MI and SMA (Figure 2B).7,10,14,16 Neurons responding to the orofacial, forelimb, and hindlimb areas of the MI are located along the ventral-to-dorsal axis in the GPe and GPi (MI domain). Neurons responding to these areas of the SMA are also located along the ventral-to-dorsal axis, but in more rostral and dorsal parts of the GPe and GPi (SMA domain). These somatotopic representations in the MI and SMA domains are partly fused in the rostro-caudal central zone.
Somatotopic Organization and Pathophysiology of Movement Disorders
- Malfunctions of the basal ganglia cause movement disorders such as Parkinson disease, Huntington disease, and dystonia. Some symptoms of these diseases can be explained by the “firing rate model,” which means that the activity imbalance between the direct and indirect pathways changes the mean firing rate of the output nuclei of the basal ganglia and induces hypokinetic or hyperkinetic disorders. For example, in parkinsonian state, dopamine depletion reduces tonic excitation of the striatal neurons in the direct pathway through the D1Rs, and tonic inhibition of the striatal neurons in the indirect pathway through the D2Rs.25,26 These changes in the direct and indirect pathways induce increased activity of the GPi/SNr neurons and decreased activity of thalamic and cortical neurons, which ultimately result in akinesia.
- Contrary to the firing rate model, recent studies have focused on the possibility that abnormal bursting and oscillatory activity in the basal ganglia play a causal role in the pathophysiology of Parkinson disease.27–30 Oscillatory activity and neuronal synchronization in the GPe, GPi, and STN in parkinsonian animals and patients include two major frequency bands: the frequency of resting tremor (4–9 Hz) and the beta band (10–30 Hz). Beta band oscillation may contribute to akinesia, as some reviews have mentioned.29,30
- In addition to these reports on the changes in the firing rates and patterns in the basal ganglia, changes in the somatotopy have been reported in movement disorders. In the normal state, although GPe and GPi neurons respond specifically to the unidirectional movement of a single joint, in parkinsonian monkeys, the GPe and GPi neurons responded to multiple movements of multiple joints, sometimes of the upper and lower limbs and of both the sides.31 In addition, in patients with Parkinson disease, neurons that respond to movements of multiple body parts were observed in the GPi and STN.32 It is considered that loss of dopamine may induce a cross talk of information related to different body parts.27
- In patients of dystonia, it has been reported that there were widened somatosensory receptive fields in pallidal neurons.33 Somatotopic disorganization was observed not only in patients but also in transgenic mice generated as a model of DYT1 dystonia.34 In these mice, many GPe and GPi neurons received convergent inputs from both the forelimb and orofacial regions in the motor cortex. This somatotopic disorganization may explain a phenomenon known as “motor overflow” in dystonic patients. When patients try to move one body part, a part of the area in the GPi which are related to another body part such could be inhibited because of the somatotopic disorganization. This may lead to unintended movements along with the intended movements.
Conclusions
- Despite these recent enormous findings about the basal ganglia and their malfunctions, we are still unable to completely explain the pathophysiology of movement disorders. For example, even with regard to Parkinson disease, it is difficult to understand the mechanism of the rigidity by using the present models. The mechanism of parkinsonian tremor also remains unsolved. Moreover, although deep brain stimulation is being widely used as a treatment of movement disorders, the mechanism of its effectiveness is still unclear. To solve these problems, we should re-visit the physiology of the basal ganglia from the viewpoint of both, basic science and clinical research.
Notes
-
The author has no financial conflicts of interest.
Figure 1Neuronal responses in each nucleus in the basal ganglia to electric cortical stimulation. A: A PSTH of a phasically active neuron in the putamen responding to MI stimulation. B: A PSTH of an STN neuron to MI stimulation. C: A PSTH of a GPe neuron to SMA stimulation. D: A PSTH of a GPi neuron to SMA stimulation. PSTH: peri-stimulus time histogram, MI: primary motor cortex, STN: subthalamic nucleus, GPi: internal segment of the globus pallidus, GPe: external segment of the globus pallidus, SMA: supplementary motor area.
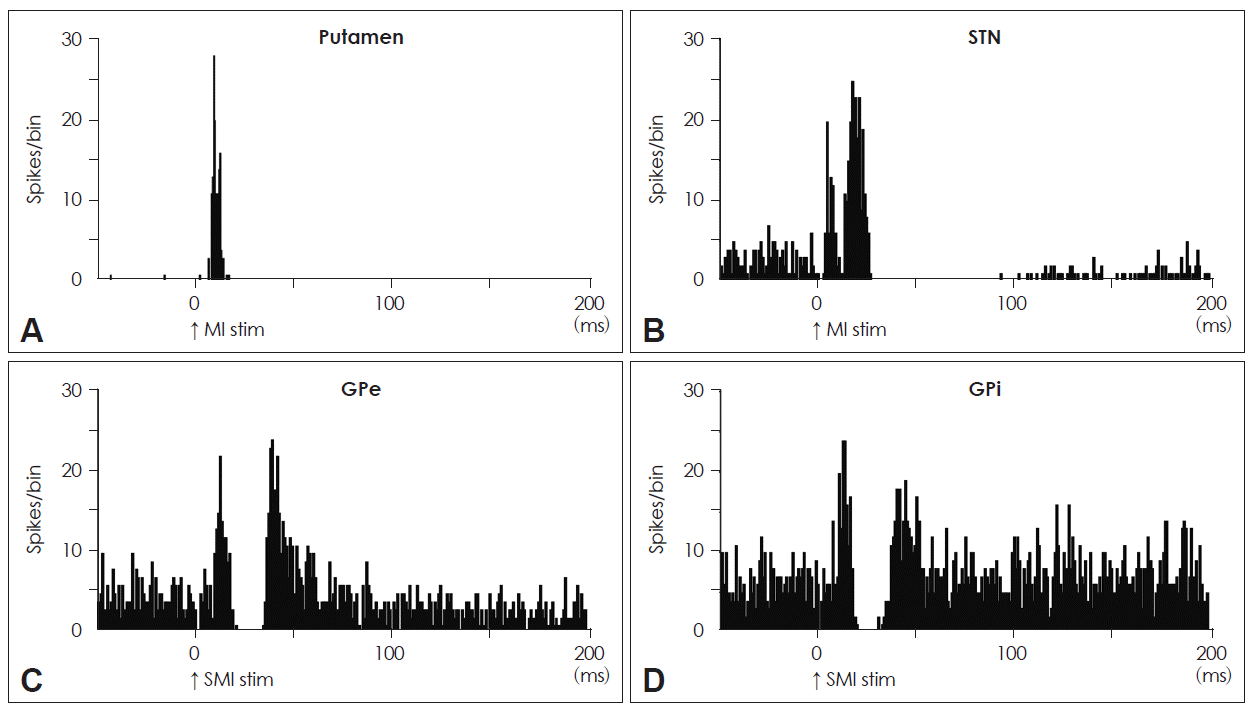
Figure 2Somatotopy of each nucleus in the basal ganglia. A: Somatotopy of the STN is shown in a frontal section. B: Somatotopic representations in the SMA domains of the GPe and GPi are shown rostrodorsally to those in the MI domains, respectively. C: Somatotopy of the putamen is shown in a frontal section. STN: subthalamic nucleus, GPi: internal segment of the globus pallidus, GPe: external segment of the globus pallidus, SMA: supplementary motor area, MI: primary motor cortex.
REFERENCES
- 1. Alexander GE, DeLong MR, Strick PL. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu Rev Neurosci 1986;9:357–381.ArticlePubMed
- 2. Alexander GE, Crutcher MD. Functional architecture of basal ganglia circuits: neural substrates of parallel processing. Trends Neurosci 1990;13:266–271.ArticlePubMed
- 3. Surmeier DJ, Ding J, Day M, Wang Z, Shen W. D1 and D2 dopamine-receptor modulation of striatal glutamatergic signaling in striatal medium spiny neurons. Trends Neurosci 2007;30:228–235.ArticlePubMed
- 4. Monakow KH, Akert K, Künzle H. Projections of the precentral motor cortex and other cortical areas of the frontal lobe to the subthalamic nucleus in the monkey. Exp Brain Res 1978;33:395–403.ArticlePubMed
- 5. Nambu A, Takada M, Inase M, Tokuno H. Dual somatotopical representations in the primate subthalamic nucleus: evidence for ordered but reversed body-map transformations from the primary motor cortex and the supplementary motor area. J Neurosci 1996;16:2671–2683.ArticlePubMedPMC
- 6. Nambu A, Tokuno H, Inase M, Takada M. Corticosubthalamic input zones from forelimb representations of the dorsal and ventral divisions of the premotor cortex in the macaque monkey: comparison with the input zones from the primary motor cortex and the supplementary motor area. Neurosci Lett 1997;239:13–16.ArticlePubMed
- 7. Hoover JE, Strick PL. The organization of cerebellar and basal ganglia outputs to primary motor cortex as revealed by retrograde transneuronal transport of herpes simplex virus type 1. J Neurosci 1999;19:1446–1463.ArticlePubMedPMC
- 8. Middleton FA, Strick PL. Basal ganglia and cerebellar loops: motor and cognitive circuits. Brain Res Brain Res Rev 2000;31:236–250.ArticlePubMed
- 9. Miyachi S, Lu X, Imanishi M, Sawada K, Nambu A, Takada M. Somatotopically arranged inputs from putamen and subthalamic nucleus to primary motor cortex. Neurosci Res 2006;56:300–308.ArticlePubMed
- 10. Akkal D, Dum RP, Strick PL. Supplementary motor area and presupplementary motor area: targets of basal ganglia and cerebellar output. J Neurosci 2007;27:10659–10673.ArticlePubMedPMC
- 11. Nambu A, Kaneda K, Tokuno H, Takada M. Organization of corticostriatal motor inputs in monkey putamen. J Neurophysiol 2002;88:1830–1842.ArticlePubMed
- 12. Nambu A, Tokuno H, Hamada I, Kita H, Imanishi M, Akazawa T, et al. Excitatory cortical inputs to pallidal neurons via the subthalamic nucleus in the monkey. J Neurophysiol 2000;84:289–300.ArticlePubMed
- 13. Iwamuro H, Tachibana Y, Saito N, Nambu A. Organization of motor cortical inputs to the subthalamic nucleus in the monkey. In: Groenewegen HJ, Berendse HW, Cools AR, Voorn P, Mulder AB, editors. The Basal Ganglia IX, Advances in Behavioral Biology. 58:New York: Springer; 2009:109–117.Article
- 14. Iwamuro H, Tachibana Y, Saito N, Nambu A. Organization of motor cortical inputs in the globus pallidus via the subthalamic nucleus in monkeys (abstr.). The 31st Annual Meeting of the Japan Neuroscience Society; 2008; P3-i20.
- 15. Nambu A, Yoshida S, Jinnai K. Discharge patterns of pallidal neurons with input from various cortical areas during movement in the monkey. Brain Res 1990;519:183–191.ArticlePubMed
- 16. Yoshida S, Nambu A, Jinnai K. The distribution of the globus pallidus neurons with input from various cortical areas in the monkey. Brain Res 1993;611:170–174.ArticlePubMed
- 17. Kita H, Nambu A, Kaneda K, Tachibana Y, Takada M. Role of ionotropic glutamatergic and GABAergic inputs on the firing activity of neurons in the external pallidum in awake monkeys. J Neurophysiol 2004;92:3069–3084.ArticlePubMed
- 18. Kita H, Tachibana Y, Nambu A, Chiken S. Balance of monosynaptic excitatory and disynaptic inhibitory responses of the globus pallidus induced after stimulation of the subthalamic nucleus in the monkey. J Neurosci 2005;25:8611–8619.ArticlePubMedPMC
- 19. Kita H, Chiken S, Tachibana Y, Nambu A. Origins of GABA(A) and GABA(B) receptor-mediated responses of globus pallidus induced after stimulation of the putamen in the monkey. J Neurosci 2006;26:6554–6562.ArticlePubMedPMC
- 20. Tachibana Y, Kita H, Chiken S, Takada M, Nambu A. Motor cortical control of internal pallidal activity through glutamatergic and GABAergic inputs in awake monkeys. Eur J Neurosci 2008;27:238–253.ArticlePubMed
- 21. Takada M, Tokuno H, Nambu A, Inase M. Corticostriatal projections from the somatic motor areas of the frontal cortex in the macaque monkey: segregation versus overlap of input zones from the primary motor cortex, the supplementary motor area, and the premotor cortex. Exp Brain Res 1998;120:114–128.ArticlePubMed
- 22. Takada M, Tokuno H, Hamada I, Inase M, Ito Y, Imanishi M, et al. Organization of inputs from cingulate motor areas to basal ganglia in macaque monkey. Eur J Neurosci 2001;14:1633–1650.ArticlePubMed
- 23. Kaneda K, Nambu A, Tokuno H, Takada M. Differential processing patterns of motor information via striatopallidal and striatonigral projections. J Neurophysiol 2002;88:1420–1432.ArticlePubMed
- 24. Smith Y, Parent A. Differential connections of caudate nucleus and putamen in the squirrel monkey (Saimiri sciureus). Neuroscience 1986;18:347–371.ArticlePubMed
- 25. DeLong MR. Primate models of movement disorders of basal ganglia origine. Trends Neurosci 1990;13:281–285.ArticlePubMed
- 26. Mallet N, Ballion B, Le Moine C, Gonon F. Cortical inputs and GABA interneurons imbalance projection neurons in the striatum of the parkinsonian rats. J Neurosci 2006;26:3875–3884.ArticlePubMedPMC
- 27. Bergman H, Feingold A, Nini A, Raz A, Slovin H, Abeles M, et al. Physiological aspects of information processing in the basal ganglia of normal and parkinsonian primates. Trends Neurosci 1998;21:32–38.ArticlePubMed
- 28. Levy R, Hutchison WD, Lozano AM, Dostrovsky JO. High-frequency synchronization of neuronal activity in the subthalamic nucleus of parkinsonian patients with limb tremor. J Neurosci 2000;20:7766–7775.ArticlePubMedPMC
- 29. Brown P. Abnormal oscillatory synchronisation in the motor system leads to impaired movement. Curr Opin Neurobiol 2007;17:656–664.ArticlePubMed
- 30. Rivlin-Etzion M, Marmor O, Heimer G, Raz A, Nini A, Bergman H. Basal ganglia oscillations and pathophysiology of movement disorders. Curr Opin Neurobiol 2006;16:629–637.ArticlePubMed
- 31. Filion M, Tremblay L, Bédard PJ. Abnormal influences of passive limb movement on the activity of globus pallidus neurons in parkinsonian monkeys. Brain Res 1988;444:165–176.ArticlePubMed
- 32. Iwamuro H, Taniguchi M, Takahashi H, Terao T, Kawasaki T, Yokochi F, et al. Receptive fields of motor-related neurons in the STN and GPi in patients with Parkinson disease. Func Neurosurg [Japanese] 2006;45:138–143.
- 33. Vitek JL, Chockkan V, Zhang JY, Kaneoke Y, Evatt M, DeLong MR, et al. Neuronal activity in the basal ganglia in patients with generalized dystonia and hemiballismus. Ann Neurol 1999;46:22–35.ArticlePubMed
- 34. Chiken S, Shashidharan P, Nambu A. Cortically evoked long-lasting inhibition of pallidal neurons in a transgenic mouse model of dystonia. J Neurosci 2008;28:13967–13977.ArticlePubMedPMC
Citations
Citations to this article as recorded by

- Parvalbumin+and Npas1+Pallidal Neurons Have Distinct Circuit Topology and Function
Arin Pamukcu, Qiaoling Cui, Harry S. Xenias, Brianna L. Berceau, Elizabeth C. Augustine, Isabel Fan, Saivasudha Chalasani, Adam W. Hantman, Talia N. Lerner, Simina M. Boca, C. Savio Chan
The Journal of Neuroscience.2020; 40(41): 7855. CrossRef
 E-submission
E-submission

 PubReader
PubReader ePub Link
ePub Link Cite
Cite